Wavegenetics device
The matrix Gariaev are produced with the help of a laser beam reflected from the object which is supposed to...


Linguistic Wave Genetics Forum

different devices working on principles wave genetics

Reviews received treatment Garyaeva matrices
our group in the social network
our group in the social network

Our Group in the social network vkontakte
The matrix Gariaev are produced with the help of a laser beam reflected from the object which is supposed to...
For the first time in the history of medicine, Doctor Nadezhda Ustinova reports full recovery in the case of “incurable”...
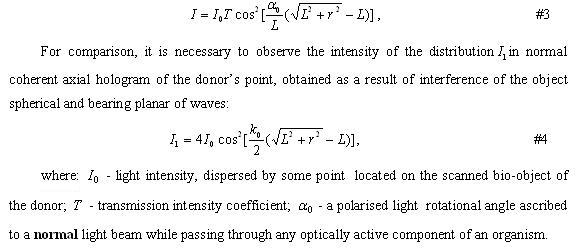
For the purpose of acquiring an insight into principles of operation of the laser holographic-data converter, which is in fact...
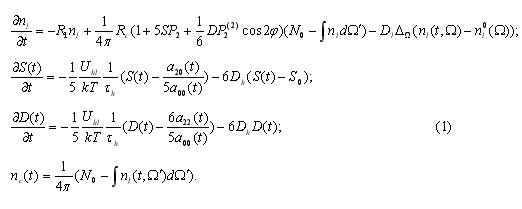
Wave Genetic Bio-Management nanotechnologies. Theory and empirical evaluation Tertishnii G.G. and Gariaev P.P. New medicine technologies, №7, p. 49-64 (2007)...
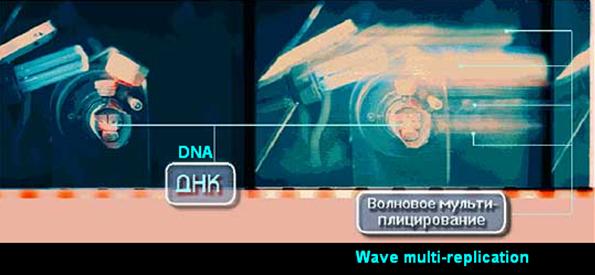
Experimental examination/investigation in vitro of holographic mapping and holographic transposition of DNA in conjunction with the information/data pool encircling DNA...

We have noted earlier (Gariaev, Maslova et all, 1996 (a); Gariaev, Maslova et all., 1996 (b)), that in the role...
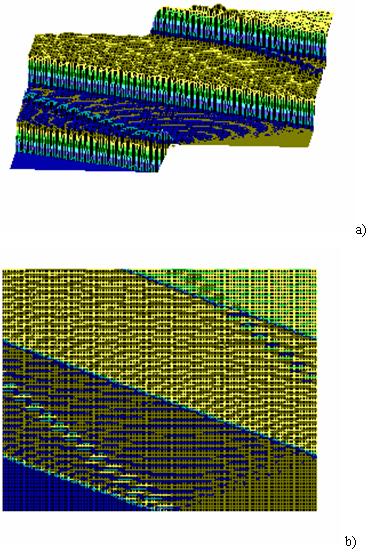
There exists another complication as well in relation to selfadjustment/coordination of biosolitons and electromagnetic wave of excessive radiation. Let’s remind...

Numerical integration of system (2) showed that two separate/solitary waves are formed, moving from the right to the left along...
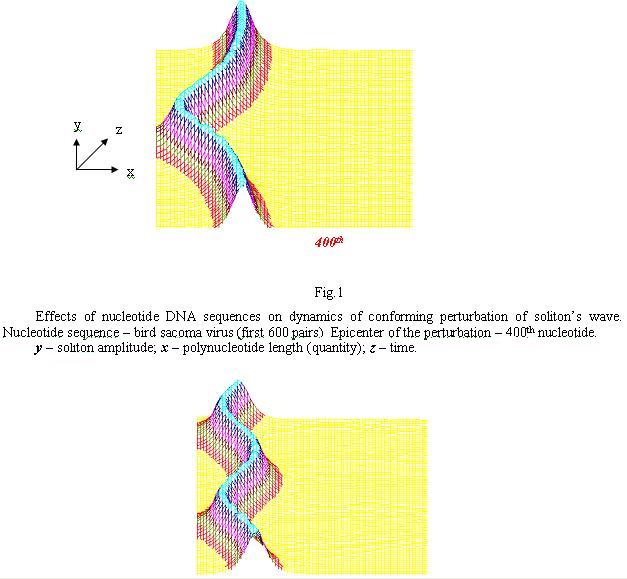
Mario Salerno was the first to experiment with solitons on DNA on computer not only as formal mathematical structures but...