Gariaev P.P., Tertishni G.G., Tovmash A.V.
Experimental examination/investigation in vitro of holographic mapping and holographic transposition of DNA in conjunction with the information/data
pool encircling DNA .
New Medical Technologies, #9, pp 42-53 (2007)
In this paper we expound the concept that irradiating DNA samples in vitro by a certain set of electromagnetic fields/frequencies in the UV and IR light spectrum causes/triggers a reaction/response in the form of protean-dynamic-long-lasting replicating holographic DNA images and its immediate surroundings.
We have discovered that DNA samples are capable of in vitro producing multiple replicating responses-images/structures, replicas of DNA itself in space and of its nearing environment. Such replicas are, in fact, responses to certain defined sets of electromagnetic fields/frequencies radiation in UV-IR range. These replicas are built up precisely horizontally to the both opposite sides, move in complex trajectories and also remain intact for a certain period of time after discontinuing the irradiation. [Refer to Fig.t.1-4.]
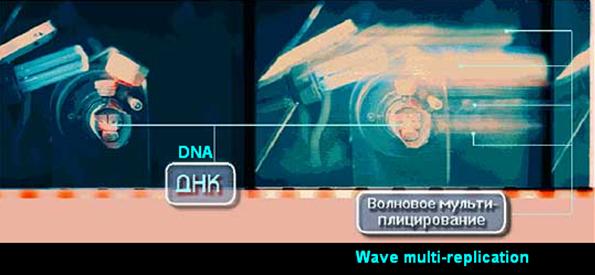
Fig. 1. Visible effect of the wave holographic replications of DNA and its immediate environment (of the objects). Control to the left and the experiment to the right. This is the second method of elicitation of DNA wave replicas.

Fig. 2 (a). Discreteness and intricacy of the trajectory of DNA wave replications (to the left – the original image, to the right — contrasted). This is the first method of elicitation of DNA wave replicas.

Fig. 2 (б). Distribution of the image brightness by RGB color model, Red, Green, Blue
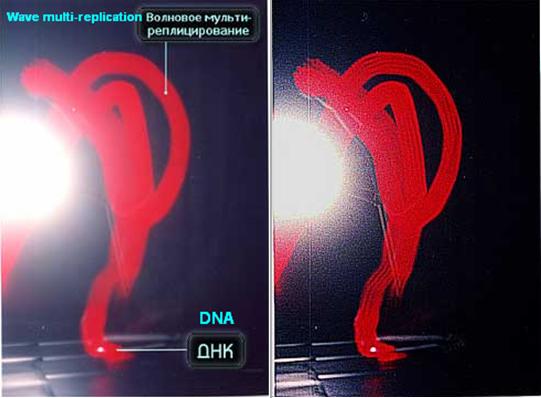
Fig. 3 (a). Spatial dynamics of DNA wave replicas. The thin (fine) longitudinal structure of the track can be observed.

Fig. 3 (б). Distribution of the image brightness by RGB color model, Red, Green, Blue.
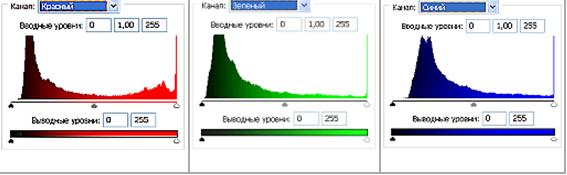
Fig 3.2. Histograms of the brightness values per RGB color model, Red, Green, Blue.
In the control photos-pictures, when the sources of the UV-IR electromagnetic fields/frequencies are turned off, this effect cannot be observed; exactly the same absence of the replicas/effects is observed when there are no DNA samples. Within the exposure time of 1 second (Refer to Methods section below) in the first version of the experiments the wave replicas move in space and multiplicate, their trajectory or path is not predictable and is discrete. Reproduction of the replicas occurs only under the condition where the camera, DNA sample and the source of the electromagnetic waves are motionless, stationary in relation to one another. An external interference with DNA sample (the second version of the experiment) engenders a change of the distribution (replica propagation) vector in space to the exact opposite (from the left to the right) and then – disappearance (dissipation). (Fig. 5 (а, б, в)). Significantly, that on a number of occasions a replication of not only of the DNA samples but of the surrounding object, takes place. (Fig. 1, 5 (a, б))
In order to analyse the color imprints (impression) on the emulsion of the film pictures are shown, which were obtained by way of scanning of the film as a distribution of brightness in RGD color mode. (Fig. 2(б), 3(б), 4(б)). In the Fig. 2 (а, б) it is visible that virtually entire brightness scale of the replicas’ image is located in Red spectrum whereas they are hardly distinguishable in Green and Blue color channels. Similar pattern of prevailing Red spectrum can be clearly observed in the accompanying Fig. 3 (б) histograms of brightness (Рис. 3 (в)) of the respective channels. Notably, one part of the histogram draws one’s attention to where replicated image is located: a sector of brightness from value of 128 to 255 – distinctive peak values of Red in that part of the chart compared to the steady decline in the other charts of Green and Blue in the same area.
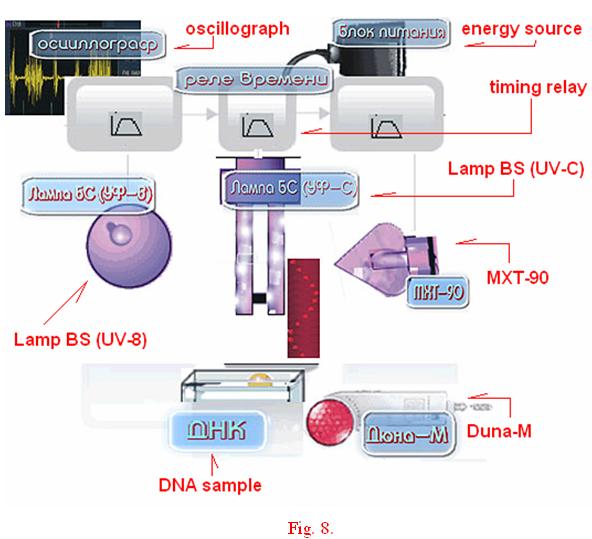
Perhaps such an apportionment of brightness may denote the fact that the photographed discrete track is in fact a pattern/replica of the light matrix of Duna-M equipment (red and InfraRed color – position 8, Fig. 8) which contained the DNA sample itself — indirectly influenced or “filtered” by the DNA sample and subsequently replicated. This equipment, this source of light is in fact a matrix of 37 alternation diodes, where out of 37 – there are 21 red and 16 infrared. The DNA sample had been placed in a test tube called эппендорф??? and was covering 5 diodes. This precisely correlates to the 5 fine longitudinal structures of the track in the replica. (Fig. 3a)
An important property of DNA wave replicas is their relatively long lifetime after turning off of all initiating electromagnetic fields/frequencies sources. (Fig. 4), however the lifetime of the replicas may also depend upon sensitivity of the photographic film, which is being used to record the replicas and also the film’s spectral sensitivity/selectiveness.
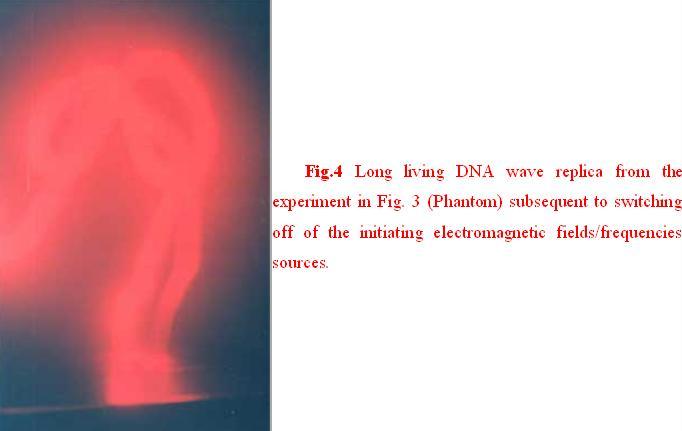
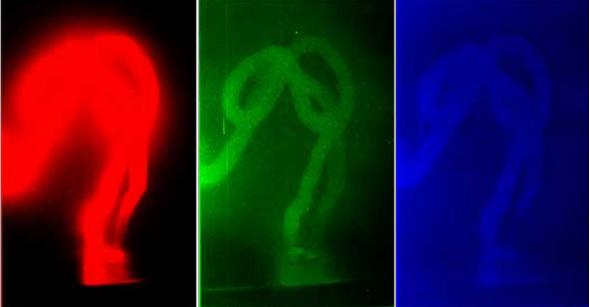
Fig. 4 (б). Distribution of the brightness values per RGB color model, Red, Green, Blue
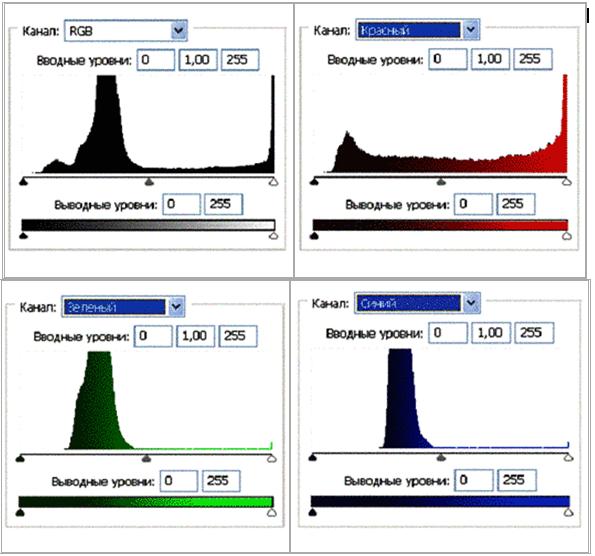
Fig. 4 (в). Histograms of the brightness values per RGB color model, Red, Green, Blue.
Notable difference is observed in the apportionment of brightness per RGB color mode between the image of the wave replica of DNA sample and its “phantom”, which remains even after the source of light is turned off. Analysis of the images in Fig. 4 (б) and juxtaposing it with the histogram in Fig. 4 (в) reveals that the image of the replica’s phantom in Red spectrum becomes blurred in it diapason as well as in its amplitude of the brightness values. That being so, Green and Blue spectrums reflect narrowing of values in the range from 70 to 120, a distinct peak is viewed which explains the brightness of the DNA replica, its variations of color and shades particularly in its Green spectrum/channel.


The type of occurring replicas in each particular case is, having regard to the methods utilized, incidental (The First method) or determined (The Second method) in space-time.
One of the outcomes of inducing wave replicas of the surrounding objects by means of DNA sample via first method, was multiplicate triplet imaging of the BS (UV-B) lamp/tube

As one of the controls to examine the capability of generating wave replicas we used sodium chloride (NaCl, crystallised); sodium chloride (1M water solution); vine acid crystallized; vine acid рацемат? (1M water solution); dry starch; glycine crystallized; dry calciferol; dry tocopherol; dry chlorophyll, bi-distilled water; interferon mixed with Bacillus subtilis – dry.
None of the mentioned substances produced any wave replicas.

One of the modified experiments, shown in Fig. 5 (old DNA sample replaced by new). Refer to the shots # 3 and #4 above. The shot #4 reveals replicas of the “Duna-M” diodes, shifting to the right side. It is tyFig.al to observe the following- appearance of replicas of perforation and exposed parts of the film close to diodes.

Shots #11 and #12 above. It is to be mentioned that from shot #1 until shot #11 the Duna-M diodes wave replicas are absent, however appearing again in the shot #12.

Shots # 13 and #14 above. In the #13 we can distinguish wave replicas of the Duna-M diodes with the commonly observed intrusion in into the unexpected space between the shots. The shot #14 does not capture the wave replicas of the diodes and they disappear.

Shots # 23 and #24 above. From shot #14 until #22 replicas disappear, yes dimly captured in the shots #23 and #24.

We have utilized two schemes developed for our experimentlal purposes. The first is produces in Fig.9, the second can simply be viewed directly in Fig. 5(в). The schemes are also given in Reference #18 in the Bibliography. In order to adduce and visualize DNA wave replicas in accordance with the first scheme, the following is to be performed: by means of timing relay (Fig. 8, position #3) in varied combinations the operator switches on the required emitters BS (UV-B, which is incandescent lamp in blue spectrum «ЛОН БС 75», glass type — СЛ98-1 – Fig 8, position #5), a matrix with red and infrared diodes (Fig.8 position #8.) and germicidal mercurial lamp/bulb or lamp Compact electronic CEST26E27 Black (UV-C, Fig 8, position #6), or BS (UV-B) and MXT-90 (cold cathode – Fig.8 position #4).
Dehydrated/dry DNA sample from bull’s spleen — about 100 milligrams in a sealed plastic conical test tube «эппендорф » 4 centimeters long, 0.9 cm in its upper end; OR 3 milliliters of DNA water solution, 1mg/ml) are placed in the effective zone of the emitters (РЕАХИМ, марка А, НПО «Биолар» ) (1mm-50cm from the light emitters) and then the emitters are activated. The progression of the experiment was filmed using Fuji 24-27 DIN film. Oscillograph is in operation during the experiment and is to register and record the electromagnetic fields/frequencies within the zone of the experiment; averaged normal electromagnetic background noise from within the premises is recorded, defined by the behavior of sinusoid in the oscillograph. Further, by means of the timing relay the emitter UV-C is turned off 10 minutes later. The camera captures an emergence of unique dynamic wave structures – multi-replicated DNA replicas and of the surrounding objects invisible to the naked eye, yet perceivable by the camera and fixed on the film. These are directly related to the photonic influences effected by the emitters on the DNA samples. In other words, multiplication of a number of reflection of DNA samples occurs and is redistributed in space on complex trajectory patterns (the first method) and on horizontal patterns (the second method), including mapping of the objects responsible for exciting the DNA samples.
The Second method to obtain and visualize DNA wave replicas involves the following: dehydrated/dry DNA sample, 100 milligrams is placed in an open mode into a holder made of aluminum foil. With intervals of 2-3 seconds the BS (UV-B) lamp, Compact Electronic CEST26E27 Black lamp and apparatus Duna-M are turned on and off. Photographs are taken 5 minutes later. By this method the DNA replicas and of close objects are registered and they are propagating strictly to the right hand side. With external mechanical interference (a touch ) with the DNA samples the distribution vector alters to the opposite – that is the DNA replicas begin propagating to the left hand side and than 5-8 seconds after the mechanical interference the replicas disappear or are not perceived by the present equipment or the utilized film, regardless of the equipment still being in an activated state.
DISCUSSION
Despite the successes of molecular genetics and cellular biology, thus far the finest mechanisms of the symbolic functions of genetic apparatus are not understood and remain to be discovered and researched. This view had most strikingly been upheld and propounded after publication of Pruitt led research team where he elegantly proven that Mendel’s Laws in genetics do not always work, moreover behavior of one of the genes does not comply with commonly accepted views. (Reference #1). This is reflected in that in adult species of Arabidopsis phenotypically emerged normal gene Hothead, which was not present in the original mutated seeds (the seeds were mutated on Hothead so as to remove it )
A view was put forward that normal gene had been stored/preserved as its revertible RNA copy. It is feeble and unsubstantiated explanation having no experimental proof. Inexplicable, from the viewpoint of classical genetics, phenomena of return/recovery of the ancestral gene brought to the surface a whole chain of unresolved cardinal issues of genetics and embryology. The issues can be briefly enumerated:
a) wobbling of the 3rd nucleotide in codon makes the canonical table of the genetic code an arena of potential errors in synthesis of proteins because it automatically gives омонимии?? important doubles/ дублетов in codons, when the first pair of identical doubles/ дублетов performs coding of different amino-acids2 and with this the third nucleotide of codon can be any of the four, that is what was postulated by F. Crick3 , nonetheless errors upon selection of amino-acids are highly unlikely and this is surprising and extraordinary fact;
b) situation where 98% of the genome eukaryote is considered to be “useless”/”garbage”, or at most – an assistant of the triplet code, or a “graveyard” of virus genomes;
c) 2% of the human coding DNA (about 40000 genes) appeares to be rather similar to the one of pigs, donkeys, flies and even E.coli;
d) genes, for some obscure reasons and laws not know to the present science, transposition in to 3-dimentional continuum of chromosomes;
e) genes are divided into Introns and Exons, for incomprehensible reasons.
f) independent matrix synthesis of RNA sequences Qβ replica/репликазой bacteriophage of E. coli and an alike synthesis of RNA sequences polymerase bacteriophage T7 E. coli4,5 which violate the canonical principle of purely material/substantial replication DNA↔RNA;
g) leaf’s phantom effect, where a part/section of a living leaf of a plant reproduces its own whole image in gas-discharge visualization (Kirlian’s equipment) 6,7;
h) Inexplicable in its mechanics/process, distant functions of selective genes of morphogenesis, where their products are synthesized in one locality, yet the action is instantly detectable/viewable at some other place, away from the developing embryo.
All these issues and others that are little understood phenomena of symbolic chromosome functions manifestation and compel one to consider and prove the fact that genetic memory possesses other, unknown, undiscovered qualities and aspects significantly complementing the protein code. Feasibly, these properties have their origins in waves/electromagnetism . Precisely these very contradictions and a lack of structured model for the triplet protein code for expounding of coding of the space-time continuum of the complex multicellular bio-systems compelled us to draw our attention to the workings of the genetic apparatus as a system of highly-advanced and highly organised symbolic emanations of electromagnetic and acoustic fields/frequencies/waves.8,9,10,11 Probably, one of the manifestations of the wave symbolic functions of DNA are the results that are being discussed in this paper – the findings, outcomes of our research into DNA wave replicas .
The very first evidential proofs of DNA being able to maintain wave/field type of memory were obtained by us in 1985. We discovered the phenomena of so called DNA phantom effect and amply in detail reported the finding in our book8. It is perhaps true that the mentioned effect is directly connected to the related findings presented in this paper. DNA phantom effect develops as follows. The method used is called dynamic laser light-dispersion (DLLD ). DNA is considered to nee a linear polymer. Spectrometer registers in its working chamber some form of hypothetical structures (objects), peculiarly dispersing the light after removing DNA samples from its working chamber. This particular fact/phenomena had been named ‘DNA phantom effect’. Test measurements, prior to placing the DNA in to the spectrometer’s chamber, register mere common background (noise) light-dispersion.
Insufflations of the spectrometer’s chamber with nitrogen gas results in disappearance of the phantom effect, however it is newly detected 5-7 minutes later. As a matter of fact similar effect had been observed by Allison et.al.13 who also used DLLD method to study restrictive fragments of DNA. The research group also observed anomalous light-dispersion/scattering of such objects. The authors defined the phenomena as ‘mimicking the effect of dust’. In their experiments the effect was not due to any form of dust contamination but was a consequence of behavior of the “object” which acted as a dust particle scattering light. To sum up the finding, such a conduct of the particles produces characteristics of light-dispersion which do not comply with the classical models of light-dispersion for linear polymers.
There can be found a common ground between8,12,13 experiments related to the fact that DNA samples were irradiated by light in the visible spectrum — 632.8 nm and 488 nm. The same occurs by initialising DNA wave replicas in these experiments. In the formation process of DNA wave replicas, dominant function is assumed by, perhaps, the emitter whose wavelength coincides/matches with the DNA’s absorption wavelength. That is, as it seems, a source of UV light. We however cannot exclude that DNA wave replicas may assume a certain role/function during the processes of genetic wave communications among cells in ontogenesis and during adulthood state of a bio-system.
It is intrinsic for the DNA samples in vitro to have quasi genetic process: DNA by means of waves duplicates itself and the objects (equipment, articles) involved in initiating of the replication (Fig. 1-3, 5 (a), (b), 6 (a), (b)),. That aspect of the conduct of the genetic apparatus in vivo in multicellular bio-systems is one of the key features of our model of wave genome8,9,10,11. In accordance with that model every single cell and the entire bio-organism effects uninterrupted scanning/examination/monitoring of its own genetic-metabolic state.
That is to say that DNA, by means of its own coherent irradiations in vivo using polarized-holographic means,10 scans itself and its internal cellular metabolic space/region/area in frequency range of 250-800 nm, that is – it is copying, creating wave/light replicas of its structural-functional status at any given moment in time. It is probable that the range of the spectrum is more diverse or wider; however we are technically limited in detecting and registering the said ranges of frequencies that are beyond 250-800 nm. These replicas form so called entangled spatial structure and non-locally (and instantly) unite informationally unite/unify the biosystem.20.
What follows next are common features of DNA wave replicas recorded on films:
1. From one shot to another the replicas appear and disappear. For Examples are given earlier in the text (Fig. 5 also shots # 13 #14)
2. Replicas appear in perforated openings and “light exposures” are also typical (Fig. 4)
3. Transitions of the replicas appear in between shots (that black dividing border) on the film where they are not meant to be and they even spread to the next shot yet to be used.
We may now present some preliminary explanations for these phenomena. The observed fact of image transition from one shot to another and exposed areas of the film about the perforated area can be explained by and accorder to selecting the wave-length from the wide spectrum of планарными (do not understand the meaning of this Russian word) waveguides. They originate between upper and lower ends of the thickness of the active photo-sensitive element layer on the film and also between the transparent plastic film and the photo-sensitive-emulsion of the photo-film. When multiple reflections are present and considering [light]-scattering upon the roughness of lacking quality планарными waveguides, and also when there is no focusing and overlap of “crawling” reflected images from the opposite sides of the film on one another, their mutual superimposition occurs followed by blurring. Pictures of the perforated openings in the film do not lose their sharpness due to their relative large-scale-ness. By large-scale-ness we mean incommensurability of the perforation openings’ sizes and the size of the photo-emulsion’s grains. Large and contrasted objects do not need focusing when translating the images to short distances commensurable with the size of the image itself. By that it is explained why there is an interference with the separation line between any two shots when large-scaled images of the luminescent diodes encroach on the separating line.
The effect of irregular registration of the replicas from one shot to another can be understood as:
a. Relatively prolonged UV-“pumping” (by means of laser waveguides) of the DNA sample tissues occurs with its subsequent over-irradiation once they have received critical amount of stored energy.
b. The energy emission within certain period of time, significantly shorter than “pumping” however.
c. When there is no co-occurrence between filming and the moment of stored/pumped energy emanation/”firing”, the shot does not captures DNA wave replicas nor does it register the surrounding objects; when there is co-occurrence between these two processes then replicas are registered on the film.
If a certain tuned/optimal timing is selected then we will register every emanation/“firing” of the stored/pumped energy in one continuous sequence on every shot made.
We have previously postulated that DNA in vivo – in vitro is in essence a hologram-creating/producing environment8.9. Having accepted this stance, we could believe that on the photo-sensitive area/location of DNA, as much as on collagen, holograms can be recorded artificially by means of blue and UV spectrums. In that case having used blue and UV lamps for irradiation, contemporaneous auto-recording of DNA sample on itself and parallel recording of the surrounding objects occurs onto photo-sensitive DNA’s quasi-cylindrical formations in a state of rigid gel. This is followed by scanning/reading (by means of red and IR emitters) from each of them of multiple diffractioned blurred and so distorted images of first bright and later weaker, of all subsequent orders/forms of diffractioned images, shifted in relation to one another.. This registration of the holograms in UV with consequent reconstruction in red and IR spectrum leads accordingly to degradation and blurring of the image. Blurring is caused due to their image being spatially disbanded in as much as because of their content including several orders/forms from each DNA thread/fiber. The blurring also occurs because of DNA’s own acoustic vibration/vacillation with the return effect of Fermi-Pasta-Ulama8.
It is our view that such replication of images under large amount of UV radiation, for instance, received as a result of sunburn, in the skin and nearing tissues of humans, can cause/create pathological programs of false holographic operation/management when scanned off/read by red or IR spectrum of the sunlight. This in turn, after subsequently exciding the threshold level of brightness for the reconstructed/altered holographic images, may lead to appearance of some sort of malignant tumors. This is already known for melanoma.
Aside from registering holograms and having considered large amounts of UV energy simultaneously with registration of holograms, effect of partial/discrete knocking out of electrons and partial violation of the DNA structures ensues, where the congregation of those electrons causes a development of free type of capacitative charge on the surface of DNA filaments/tissues. Thereby the accumulated charge creates an effect of spatial redistribution of DNA filaments/fibers/threads, the fact which in turn affects a predominant distribution of the reconstructed images. Shifting of the images, rebuilding the diffractioned orders, to the side opposite to the original diffraction is determined by the capacitative effect of alternating the charge to the opposite – e.g. minus to plus or plus to minus. Having come in contact with such spatially-scattered quasi-capacitor because of the fractional leakage and redistribution of the charges and their mutual location new phenomenon has been discovered of predominant appearance of right or left diffraction orderings in the controllable DNA nanostructures. This very effect can be observed in Fig. 5 (а, б). This very effect will be possible to use for creating controllable/manageable spatial DNA nanostructures, for instance, in the processes of human tissue and human organs regeneration as a result of purposeful holographic management/operation, which is already achieved in its primary version. [Reference to 21, 22]
We ought to underline/emphasise that the discovered phenomena of DNA wave replicas formation and of its immediate environmental objects requires more rigorous, precise and comprehensive experimental setting and theoretical substantiation. A lot remains uncertain and obscured and therefore the research continues.
Bibliography
1. Lolle S.J., Victor J.L., Young J.M., Pruitt R.E. Genome-wide non-mendelian inheritance of extra genomic information in Arabidopsis. Nature, 434, 505-509 (2005).
2. Lagerkvist U., «Two out of Three»: an alternative method for codon reading. Proc. Natl.Acad.Sci.USA, 75, 1759 1762 (1978).
3. Francis Crick. A personal View of Scientific Discovery. Basic Books, Inc., Publishers, New York. Translate into Russian: Institute of computer researches, Moscow-Ijevsk (2004), p. 98.
4. Biebricher C.K., Eigen M., Luce R. Product analysis of RNA Generated de novo by Qb Replicase. J.Mol.Biol., 148, 369-390 (1981).
5. Biebricher C.K., Luce R., Template-free generation of RNA species that replicate with bacteriophage T7 RNA polymerase. The EMBO Journal, 15, Issue 13, 3458-3465 (1996).
6. Choudhury G.K., Kejarival P.C., Chattopadhyay A. The devices for generation of phantom leaf effect. J.Inst.Eng. (India) 60, Pt.EL3, 61-66; 67-73 (1979).
7. Gariaev P.P., Junin A.M. Phantom leaf effect. Myth or reality? Energy 10, 46-52 (1989). In Russian.
8. Gariaev P.P. Wave based genome. Monograph. Moscow. Ed. Obshestv. Pol’za. 279p. (1994). In Russian.
9. Gariaev P.P. Wave genetic code. Monograph. Moscow. Ed. Izdatcentr. 108p. (1997). In Russian.
10. Prangishvili I.V., Gariaev P.P., G.G.Tertishny, V.V.Maximenko, A.V.Mologin, E.A.Leonova, E.R.Muldashev. Spectroscopy of radiowave radiation of localized photons: path to quantum nonlocality of bioinformation processes. Sensors and systems, 9 (18), 2-13 (2000). Russian Academy of sciences. In Russian.
11. Gariaev P.P., Birshtein B.I., Iarochenko A.M, Marcer P.J., George G. Tertishny G.G., Leonova E.A., Kaempf U., The DNA-wave biocomputer. «CASYS» – International Journal of Computing Anticipatory Systems (ed. D.M.Dubois), Liege, Belgium, 10, 290-310, (2001).
12. Gariaev P.P., Chudin V.I., Komissarov G.G., Berezin A.A., Vasiliev A.A., Hologrphic Associative Memory of Biological Systems, Proceedings SPIE — The International Society for Optical Engineering. Optical Memory and Neural Networks, 1621, 280- 291. USA. (1991).
13. Allison S.A., Sorlie S.S., Pecora R., Brownian Dynamics Simulations of Wormlike Chains, Dynamic Light Scattering from a 2311 Base Pair DNA Fragment. Macromolecules, 23, 1110-1118, (1990).
14. Hagerman, P. J., and B. H. Zimm., Monte Carlo approach to the analysis of the rotational diffusion of wormlike chains. Biopolymers, 20, 1481–1502, (1981).
15. Zimm B H., Dynamics of polymer molecules in dilute solution: viscoelasticity, flow birefringence and dielectric loss, J. Chem. Phys., 24, 269-78, (1956). General Electric Research Laboratory, Schenectady, NY.
16. Rouse P E., A theory of the linear viscoelastic properties of dilute solutions of coiling polymers, J.Chem.Phys., 21, 1272-80, (1953). Tertyshnii G.G., Gariaev P.P., Aksenov V.A., Leonova E.A., Fomchenkov S.V., The formalism of endogenous polarization/holographic managing processes in organisms. Consciousness and a physical reality, 9, number 4, 44-50 (2004). In Russian.
17. Prangishvili I.V., Gariaev P.P., Tertishnii G.G., Maksimenko V.V., Mologin A.V., Leonova E.A., Muldashev E.R., Spectroscopy of radiowave radiations of the localized photons: an output on quanum nonlocal bioinformation processes. Sensors and Systems, 9 (18), 2-13 (2000). In Russian.
18. An application for a Patent to the Federal Institute of Industrial Property of Russia, 20.10.2005. Registration number # 2005132403, Incoming number # 036308.
19. Tertyshnii G.G., Gariaev P.P., Aksenov V.A., Leonova E.A., Fomchenkov S.V., The formalism of endogenous polarization/holographic managing processes in organisms. Consciousness and a physical reality, 9, number 4, 44-50 (2004). In Russian.
20. Prangishvili I.V., Gariaev P.P., Tertishnii G.G., Maksimenko V.V., Mologin A.V., Leonova E.A., Muldashev E.R., Spectroscopy of radio wave radiations of the localized photons: an output on quanum nonlocal bioinformation processes. Sensors and Systems, (18), 2-13 (2000). In Russian.
21. Gariaev P P, Kokaya AA, Mukhina I V, Leonova-Gariaeva E A,
An effect of modulated electromagnetic radiation on the progression of alloxan diabetes in rats. Bulletin of Experimental Biology and Medicine, #2, p 155-158 (2007) . in Russian Language.
22. Артюх В.Д. – Artyukhin V.D., Гаряев П.П. — Gariaev P.P., Кокая А.А. – Kokaya A.A., Леонова-Гаряева Е.А. — Leonova-Gariaeva E A. , Мулдашев Э.Р. – Muldashev E.R., Мухина И.В. – Mikhina I.V., Смелов М.В. – Smelov M.V., Товмаш А.В. – Tovmash A.V., Чалкин С.Ф. – Chalkin S.F., Шатров Я.К. – Shatrov Y.K., Ягужинский Л.С. – Yagujinski L.S., 2007,
Effect of laser induced resistance/immunization in animals to the alloxan type diabetes. In Russian language. http://www.trinitas.ru/rus/doc/0016/001b/00161365.htm