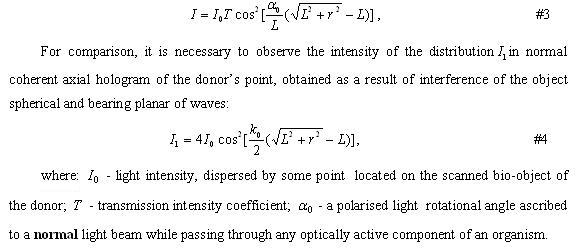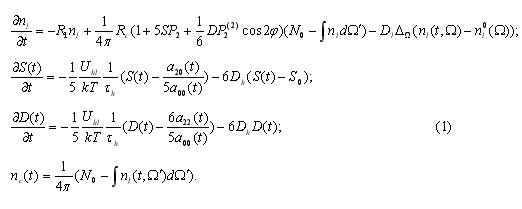Antenna Model
We have noted earlier (Gariaev, Maslova et all, 1996 (a); Gariaev, Maslova et all., 1996 (b)), that in the role of the main molecules perceiving external electromagnetic fields as a regulators – ar the proteins and especially metal containing. Functioning of a set of biological macro-moleculs (ferments in particular) is largely determined by processes occurring in the active centres surrounded by bio-polimer sequences having a sign topology. Based on this perception of informational bio-macro moleculs, it is normal to suppose that theyir interaction with physical fields external in relation to the biosystem and internal radiations leads to excitation dipolar-active fluctuations of monomers, forming the said chain, and they in turn induce fluctuation if the active centre. In toher words, such system will work as a peculiar antenna.
There excited fluctuations can lead to transitioning of bio-macr-molecul in to toher conforming (topological, sign) state.
Such conception in a principal plane is adequate to the whole set of functionally important bio-macro molecule, for example, chlorophyll, hemoglobin, mioglobin and so on,. These macromolecules are joined by two structural properties: 1) in their geometrical centre there is ion (in the case of chlorophyll – ion of magnium, in the case of hemoglobin –ion of iron); 2) near ion symmetrically placed 4 pirrolik rings (pseudo structure. )
Other types of polymers, corresponding to antenna model, can be relatively simple cycles such as valinomycine (potassium ions’ carrier) and complex over-molecular chromosom structures, DNA of which contains higly organized associates of such metals as magnesium, calcium, nickel, cobalt, copper, ferrum, zink and so on. Their role is not clear and is considered to neutralize OH-groups of remnants of phosphoric acid of polynucleotide. It is proposed that functions of metals in DNA and RNA is substantially wider and is realized along the lines of sign and/or energy interchangewith endo – and exogenous, in relation to the biosystem, physical fields. The same is applicable to the proteinsnot containing porfirine centre yet specifically binding metals, e.g. as such you can considersite-specific proteins with domens like “zink fingers” which participate in regulation genes, often located long awayfrom these governing proteins. Atoms of metals in DNA and proteins can in resonance interact via electromagnetic channels within the meaning of antenna model. Let defined the definition of antenna model.
External energy (in particular, connected with resonance interaction extremely high frequency electromagnetic radiation with proteins) travels to the periphery, that is onto ensemble of sub-units (not necessarily identical in structures). As a result of an active “conversation” predetermined by bio-chemical links between peripheral acceptor (received encoded energy) and centre-associate (in this case and ion of irongem-containing proteins), the latter receives energy (information) which effects the biological action. The degree of reactive ability of bio-macromoleculs is considerably dependent on the level of excitation of central sub-units. Lets look closely in to potential mechanisms of wave interactions of physical fields and active centres of informational bio-moleculs within the boundries of the proposed antenna model.
As an example of the simliest model for illustrating antenna effect we consider 2-dimentional closed (cyclic) chain of monomers. There is anactive centre, connected to monomers of chain by dipole-dipole interaction.






Bibliography
1. Albada P. van, Lagendijk A., 1985, Observation of Weak Localization of Light in a Random Medium, Phys. Rev. Lett. v.55, p. 2692-2695.
2. Bennet C.H., Brassard G., Crepeau C., Jossa R., Peres A., Wootters W.K., 1993, Teleporting and unknown quantum state via dual classical and Einstein-Podolsky-Rosen channels. Phys.Rev.Lett.,v.70, p.1895-1899.
3. Bouwmeester D., Pan Jian-Wei, Mattle K., Eibl M., Weinfurter H., Zeilinger A., 1997, Experimental quantum teleportation. Nature, v.390, p.575-579.
4. Einstein A., Podolsky B., Rosen N., 1935, Can quantum-mechanical description of physical reality be considered complete? Phys.Rev. v.47, p.777-780.
5. Fermi E., Pasta J., Ulam S., 1955, Studies of nonlinear problems. 1. Physics. Report. http://www.osti.gov/accomplishments/pdf/A80037041/A80037041.pdf
6. Gariaev P.P., Chudin V.I., Komissarov G.G., Berezin A.A., Vasiliev A.A., 1991, Hologrphic Associative Memory of Biological Systems, Proceedings SPIE — The International Society for Optical Engineering. Optical Memory and Neural Networks., v.1621, p.280- 291. USA.
7. Gariaev P.P., Birshtein B.I., Iarochenko A.M., Marcer P.J., Tertishny G.G., Leonova K.A., Kaempf U., 2001, The DNA-wave biocomputer. “CASYS” – International Journal of Computing Anticipatory Systems (ed. D.M.Dubois), Liege, Belgium, v.10, pp.290-310; http://www.rialian.com/rnboyd/dna-wave.doc
8. Gariaev P.P., 2003, Der wellengenetische Code. Tattva Viveka, №20, pp.68-73. (in German).
9. Lolle S.J., Victor J.L., Young J.M., Pruitt R.E. Genome-wide non-mendelian inheritance of extra genomic information in Arabidopsis. Nature, 434, 505-509 (2005).
10. Lushnikov A.A., Maksimenko V.V.,.Simonov A.J, 1982, Electromagnetic Surface Modes in Small Metallic Particles, in Electromagnetic Surface Modes, ed. by A.D.Boardman, J. Wiley, Chichester, pp. 305-345.
11. Maksimenko V.V., 1999, Antoine’s Localization of Photon inside Fractal Cluster, Fractal in Engineering, Delft, Netherlands, p. 355-358.
12. Shcherbak V.I., 2003, Arithmetic inside the universal genetic code. BioSystems, v.70, pp.187–209.
13. Александров С.А. Некогерентный метод получения голограмм. // Оптика и спектроскопия. 1998, Т.85, № 6, С. 1029-1032.
14. Березин А.В., Курочкин Ю.А., Толкачев Е.А., 2003, Кватернионы в релятивистской физике. М.: УРСС.
15. Берсукер И.Б., 1976, Электронное строение и свойства координационных соединений. Ленинград. Химия.
16. Bischof M. Biophotonen. Das Licht in unseren Zellen. Printed in Germany, 1995, ISBN 3-86150-095-7, ZWEITAUSENDEINS, S. 522.