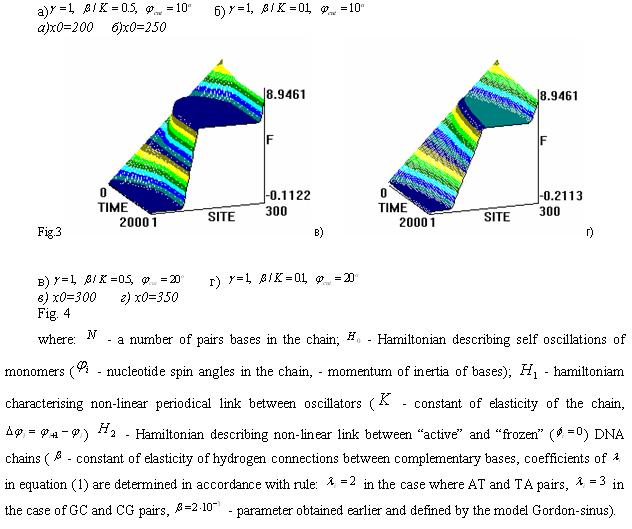
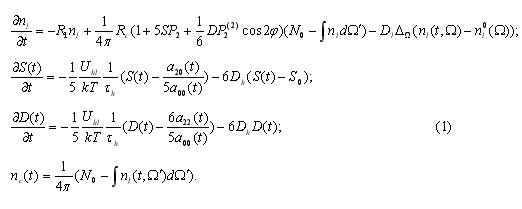Mathematical Modeling of solitons on DNA
Mario Salerno was the first to experiment with solitons on DNA on computer not only as formal mathematical structures but also tried to link their behavior in a mono-dimensional space of poli-nucleotides with their bio-genetic, or more accurately with their epi-genetic functions. He had been developing the first soliton model on DNA proposed by Englander and et al. This model and its more detailed variations including ours (see further below) is represented in term of mechanical systems as a chain of oscillators (DNA base) linked by resilient non-linear sugar-phosphate connections. Following Salerno we have paid particular attention to objectively existing known DNA sequences and thei effects onto solitons behaviour.
In the first stage we have replicated his experiments yet on significantly longer DNA parts. Indeed, solitonic kink excitations are sensitive to the place of their initiation; their motion along one of DNA chains (when they are opened as a result of temperature fluctuations) is accompanied by specific modulation of kink trajectory in time and in single-dimension space of poly-nucleotides. Such solitons are structures that emit electromagnetic and acoustic fields; their inner oscillation structures are capable of reflecting and retranslating texts and other symbolic structures of DNA in to into and out of cellular space at least at the level of large blocks of sequences. As an example behavior of kink on a fragment of singular DNA chain, which is 1020 nucleotides long from a virus of bird sarcoma
C (or S ) area of DNA (1 1020 nucleotide) on 3’-end of virus of bird sarcoma contains several “semantically” certain areas such as polypeptide-coding area (between 558 and 675 nucleotides); PolA (936) 3’-end of virus RNA, poly-adeniling site; 916 nucleotide — 5’-end of virus RNA (“capping site”); Red-sector — () – short end replica of the virus genome; Pro – possible component of observing the transcription (between 870-9000); palindrome – “pin” (870 912) .
(5’ beginning) GGC CTA TGT GGA GAG GAT GAA CTA CGT GCA CCG AGA CCT GCG GGC GGC CAA CAT CCT GGT GGG GGA GAA CCT GGT GTG CAA GGT GGC TGA CTT TGG GCT GGC ACG CCT CAT CGA GGA CAA CGA GTA CAC AGC ACG GCA AGG TGC AAG TTC CCC ATC AAG TGG AGA GCC CCC GAG GCA GCC CTC TAT GGC CGG TTC ACC ATC AAG TCG GAT GTC TGG TCC TTC GGC ATC CTG CTG ACT GAG CTG ACC ACC AAG GGC CGG GTG CCA TAC CCA GGG ATG GGC AAC GGG GAG GTG CTG GAC CGG GTG GAG AGG GGC TAC CGC ATG CCC TGC CCG CCC GAG TGC CCC GAG TCG CTG CAT GAC CTT ATG TGC CAG TGC TGG CGG AGG GAC CCT GGA GGA GCG GCC CAC TTT TCG AGC TAC CTG CAG GCC CAG CTG CTC CCT GCT TGT GTG TTG GAG GTC GCT GAG TAG TGC GCG AGT AAA ATT TAA GCT ACA ACA AGG CAA GGC TTG ACC GAC AAT TGC ATG AAG AAT CTG CTT AGG GTT AGG CGT TTT GCG CTG CTT CGC GAT GTA CGGGCC AGA TAT ACG CGT ATC TGA GGG GAC TAG GGT GTG TTT AGG CGA AAA GCG GGG CTT CGG TTG TAC GCG GTT AGG AGT CCC CTC AGG ATA TAG TAG TTT CGC TTT TGC ATA GGG AGG GGG AAA TGT AGT CTT ATG CAA TAC TCT TGT AGT CTT GCA ACA TGG TAA CGA TGA GTT AGC AAC ATA CCT TAC AAG GAG AGA AAA AGC ACC GTG CAT GCC GAT TGG TGG AAG TAA GGT GTA CGA TCG TGC CTT ATT AGG AAG GCA ACA GAC CGG GTC TGA CAT GGA TTG GAC GAA CCA CTG AAT TCC GCA TCG CAG AGA TAT TGT ATT TAA GTG CCT AGC TCG ATA CAA TAA ACG CCA TTT GAC CAT TCA CCA CAT TGG TGT GCA CCT GGG TTG ATG GCT GGA CCG TCG ATT CCC TAA CGA TTG CGA ACA CCT GAA TGA AGC AGA AGG CTT CATT 1020 (3’-ending)
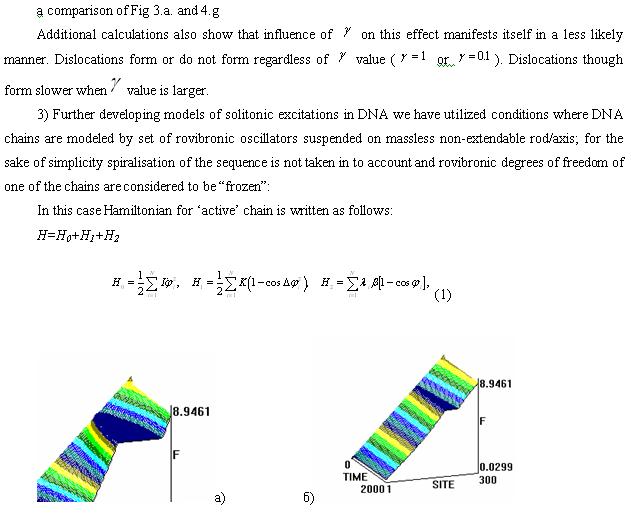
In Fig.1 and 2 the kinks appear in a form of “mountine ranges” rather tha as steps, because a derivative of sinus-Gordon equation function was taken. In here a horizontal axis – DNA sequence, vertical – soliton amplitude. Axis to the observer – time. It is visible that wnen soliton’s initiation location is altered at certain sequences of poly-nucleotide, the dynamics of that single range/wave is notably alters too in a form of spinning-fluctuating motions along DNA sequence.
The molecul’s area under this research is rich functionally (semantically) by biologically significant sectors and we are reasonably expecting that these sectors will alter, modulate, that is – will be introducing/injecting in to a single chain of DNA or RNA “textual” information. This such aspect will be realized in the spectrum of solitonic wave fluctuations alongside poly-nucleotide chain. This spectrum will mirror sequences of nucleotide and by that will perform a role of the carrier of genetic information/data/messages. Such modulation of the fluctuation structures is clearly observed in the presented graphs. It is plausible to believe that spectral composition of oscilating frequencies of solitons appear to be one of the mechanisms of text conversion of DNA and RNA structures in to wave form and a means of transmitting genetic and other messages in a mono-dimentional space alongside the poly-nucleotide chains and/or in 3D of the genome as a single cell as well as the tissue continuum of the bio-system.
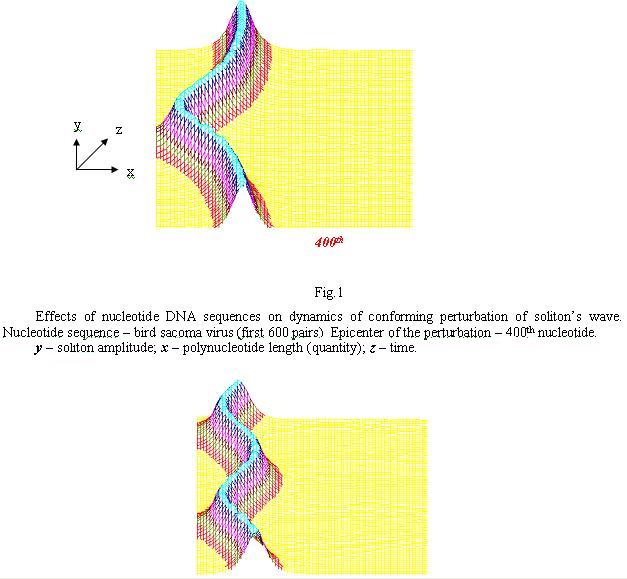
The same as Fig.1, apart from the epicenter of the perturbation – 450th nucleotide in here.
This is how the soliton dynamics computer modeling works developed by Salerno after it had been propounded by Englender. Salerno gave formalism describing spin nucleotide fluctuations of DNA molecule in order to explain experimental data in hydrogen-tritium exchange in DNA. In accordance with that model, Engleder, in DNA chain there can appear (under thermal radiation/noise) and distribute open states (“melting” the DNA double-helix in short segments enriched by AT-couples) in the form of localized dislocations.
Mario Salerno continuing works of Englender in a simplified form, discovered influence of nucleotide sequence on none-linear solitonic dynamics of spin fluctuations of nucleotides within (однотяжных- word is not known by the translator)… DNA areas, which form such open fields (open states). Later, Yakushevich, Fedianin and Khomma et al. revied various generalizations of Englender’s model assessing specifics of DNA structuring taking in to account a break in hydrogen connection when the base is opened, paired DNA chains and other aspects of reedom distinct from spinning. However, as it seems, it is not shown in those works the reasons for emergence of dislocations in DNA.
We put forward a possible mechanism of that process in DNA, which is alternative to Englender’s hypothesis about thermal noise influence as a cause for opening the bases of couples. We think that dislocations in DNA can be triggered when the period of DNA double-helix is altered (Maslov’s proposition). In our model DNA nucleotides are viewd as oscillators suspended on masless non-extendable rod/pivot; sugaf-phosphate links between neighbor nucleotides in the chain are modeled by linear springs, spiralisation along the chain is not taken in to account; hydrogen links between complementary bases are modeled by “gravitational” potential. Hamiltonian, as in accordance with M. Salerno, liiks like the following: