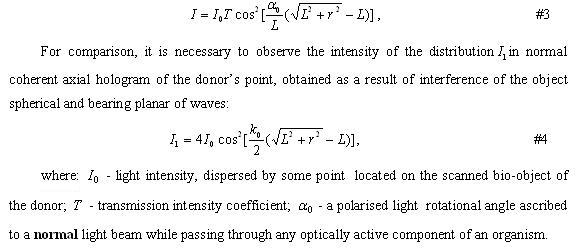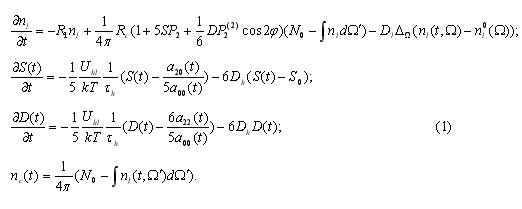PETR — ZDES 4toto BILO? Nadeyus ya ni4ego ne udalil otsuyda
There exists another complication as well in relation to selfadjustment/coordination of biosolitons and electromagnetic wave of excessive radiation. Let’s remind that mathematical modeling in this case was conducted in respect of monotonous poly-A DNA and therefore remained obscure whether heterogenic normal sequence of DNA influences the dynamics of solitonic excitation in a molecule. For the purposes of verification/confirmation , as earlier, c-aread of DNA was taken on 3’-end of sarcoma virus of birds as a field of soliton launch on various sections of the polimer. This time a derivative of of the function was calculated to show demonstratively the motion of solitons.
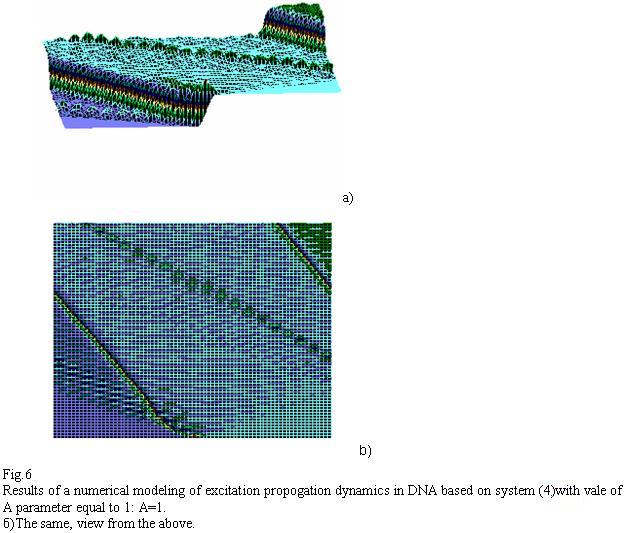
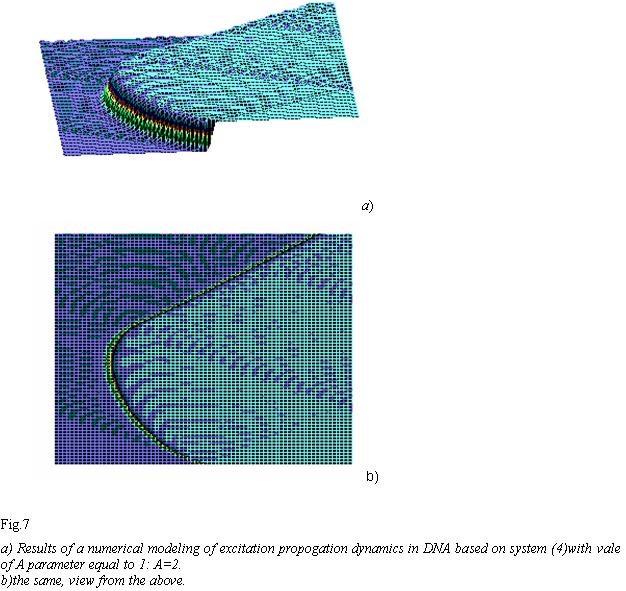
Similar to Fig. 1-3, Fig. 5-7 distinctly shows significant modifications in solitons’ behavior when altering parameter “A”. This is more evident in Fig. 7, where solitonic wave travels, similar to the one in Fig. 5-6, to the left in the beginning and then sharply turns to the right. A certain biological inference/meaning can be adduced from this fact. Soliton as a potential “reader” of DNA must “review/browse” prolonged contextual zones and not to be stuck fluctuating sinusoidally on the same “words”-locuses of DNA and RNA.
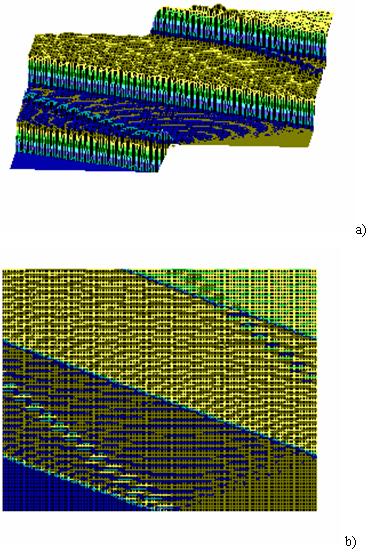
Fig.5 Results of a numerical modeling of excitation propogation dynamics in DNA based on system (2)with vale of A parameter equal to 1: A=1. The same, view from the above
If we consider non-linearity of co-valent lincs in sugar-phosphate carcass of DNA then there are further properties observable in solitons’ behavior (Fig. 8-10):
Shifting of the area of initiation ofsolitonic wave in DNA-phragment of the sarcoma virus of birds from the 200th to 500th nucleotide, then additional spinning fluctuation waves can be observed propogating to opposite directions from the base of the excitation wave. They, bouncing back from the fixed ends of DNA (in vivo the fixators’ role is taken by nucleosoms), return to the central disturbance and further modulate it. Such additional waved can be playing a role of “informers” about nucleotide structure/makeup and sequence of the bases in the area of DNA or RNA being examined, and this information can be “memorized” at the level of return of Ferni-Pasta-Ulama and be used by chromosomic biocomputer for making appropriate “decisions”
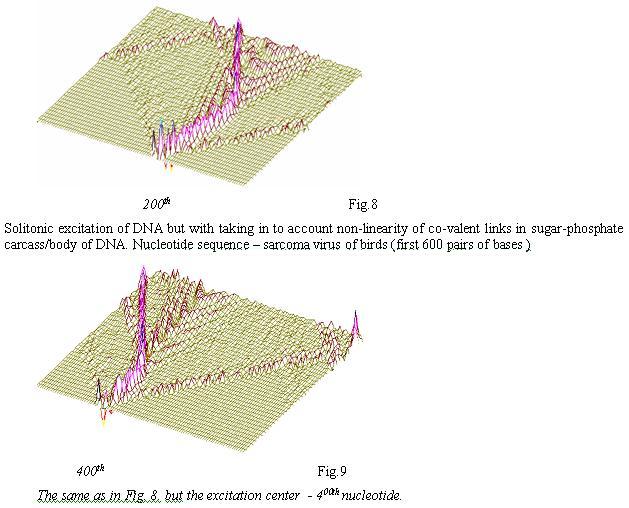
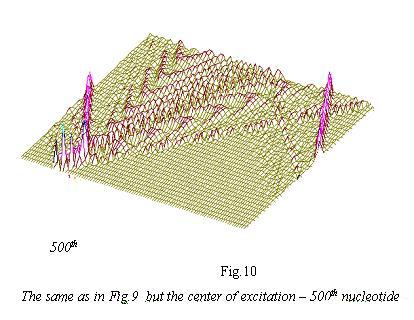
Substantial feature of solitonic DNA “scannings” can especially clearly be seen in Fig.8-10, — this is a presence of additional trajectories of solitons with rich modulation. Such additional modulated trajectories, possessing klink and briser structure, can bear additional fine nuances of redistribution of wave genetic information along the chains of DNA and RNA.